| 出品公司: | Promega |
|---|---|
| 载体名称: | pRL-null |
| 质粒类型: | 海肾荧光素酶报告载体;哺乳动物载体 |
| 高拷贝/低拷贝: | 高拷贝 |
| 克隆方法: | 限制性内切酶,多克隆位点 |
| 启动子: | CMV |
| 载体大小: | 3320 bp |
| 5' 测序引物及序列: | CMV Forward: CGCAAATGGGCGGTAGGCGTG |
| 3' 测序引物及序列: | -- |
| 载体标签: | -- |
| 载体抗性: | 氨苄青霉素 |
| 筛选标记: | -- |
| 克隆菌株: | TOP10等常规菌株 |
| 宿主细胞(系): | 哺乳动物细胞 |
| 备注: | -- |
| 产品目录号: | E2271 |
| 稳定性: | 瞬表达 或 稳表达 |
| 组成型/诱导型: | 组成型 |
| 病毒/非病毒: | 非病毒 |
pRL-null
价格:元
规格: 2ug质粒(仅用于转化提取) 转染级去内毒素液体即用质粒100μg 转染级去内毒素液体即用质粒500μg 转染级去内毒素液体即用质粒1mg 更大规格-质粒构建-质粒定制-请联系我们
联系方式:I47-825O-882O
买家导航

pRL-null载体质粒基本信息
pRL-null质粒图谱载体图谱和pRL-null载体序列质粒序列多克隆位点信息
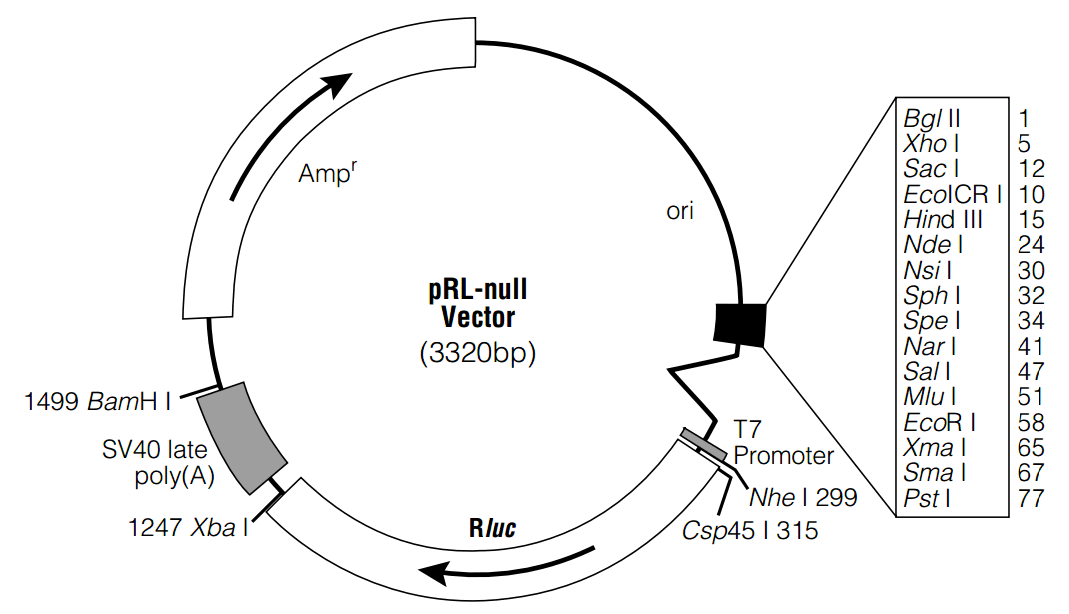


pRL-null质粒载体简介
The pRL-null Vector(a,b) (Figure 1) is intended for use in constructing a control reporter vector that may be used in combination with any experimental reporter vector to cotransfect mammalian cells. All of Promega’s pRL Reporter Vectors contain a cDNA(b) (Rluc) encoding Renilla luciferase, which was originally cloned from the marine organism Renilla reniformis (sea pansy; 1). As described below, the Renilla luciferase cDNA contained within the pRL Vectors has been modified slightly to provide greater utility. The pRL-null Vector contains no enhancer or promoter elements. Rather, it contains a multiple cloning region upstream of Rluc to allow for the cloning of any desired regulatory element(s) to drive expression of Renilla luciferase. Renilla luciferase is a 36kDa monomeric protein that does not require post-translational modification for activity (2). Therefore, like firefly luciferase, the enzyme may function as a genetic reporter immediately following translation. For information about the use of this plasmid in conjunction with a reporter vector containing the firefly luciferase gene, refer to the Dual-Luciferase Reporter Assay System(c,d) Technical Manual (#TM040). The pRL Vectors are isolated from a dam–/dcm– E. coli K host strain, allowing digestion with restriction enzymes that are sensitive to dam and dcm methylation. The GenBank/EMBL Accession Number for the pRL-null Vector is AF025844. Features of the pRL-null Vector A. Multiple Cloning Region The pRL-null Vector contains a multiple cloning region positioned immediately upstream of the chimeric intron and Renilla luciferase reporter gene (Figure 2). To aid in devising cloning strategies, Table 1 summarizes the types of DNA ends generated from restriction endonuclease digestion within the multiple cloning region as well as the compatibility of those ends with the ends of DNA fragments generated by heterologous restriction enzymes. B. Chimeric Intron Downstream of the multiple cloning region of the pRL-null Vector is a chimeric intron comprised of the 5′-donor splice site from the first intron of the human β-globin gene, and the branch and 3′-acceptor splice site from an intron preceding an immunoglobulin gene heavy chain variable region (3). The sequences of the donor and acceptor splice sites, along with the branchpoint site, have been modified to match the consensus sequences for optimal splicing (4). Transfection studies have demonstrated that the presence of an intron flanking a cDNA insert frequently increases the level of gene expression (5–8). In the pRL-null Vector the intron is positioned 5′ to Rluc to minimize the utilization of cryptic 5′-donor splice sites that may reside within the reporter gene sequence (9). C. T7 Promoter A T7 promoter is located downstream of the chimeric intron and immediately precedes the Rluc reporter gene. This T7 promoter can be used to synthesize RNA transcripts in vitro using T7 RNA Polymerase (Cat.# P2075). T7 RNA Polymerase can also be used to synthesize active Renilla luciferase in a cell-free coupled eukaryotic in vitro transcription/translation reaction (e.g., Promega’s TNT Reticulocyte Lysate(c,e,f,g) [Cat.# L4610], TNT T7 Coupled Wheat Germ Extract(c,e,f,g) [Cat.# L4140] or TNT T7 Quick Coupled Transcription/Translation (c,e,f,g,h) [Cat.# L1170] Systems). D. Renilla Luciferase Reporter Gene (Rluc) The Renilla luciferase cDNA inserted into all of the pRL Vectors is derived from the anthozoan coelentrate Renilla reniformis (1) but contains nucleotide changes that were engineered during the construction of the individual vectors. The following bases were altered in the pRL-null Vector: base 539 (T→C), to eliminate an internal Bgl II site; base 1082 (T→C), to eliminate an internal BamH I site; base 1115 (C→T), to eliminate internal Nar I, Kas I, Ban I and Acy I sites. These nucleotide substitutions do not alter the amino acid sequence of the encoded Renilla luciferase reporter enzyme. E. SV40 Late Polyadenylation Signal Polyadenylation signals cause the termination of transcription by RNA polymerase II and signal the addition of approximately 200–250 adenosine residues to the 3′-end of the RNA transcript (10). Polyadenylation has been shown to enhance RNA stability and translation (11,12). The late SV40 polyadenylation signal, which is extremely efficient and has been shown to increase the steadystate level of RNA approximately 5-fold over the early SV40 polyadenylation signal (13), has been positioned 3′ to the Rluc gene in the pRL-null Vector to increase the level of Renilla luciferase expression. IV. Transfection of Mammalian Cells with the pRL-null Vector The pRL-null Vector, once it has been modified to contain appropriate genetic regulatory domains, may be used in combination with any experimental reporter vector to cotransfect mammalian cells. However, it is important to realize that trans effects between promoters on cotransfected plasmids can potentially affect reporter gene expression (14). Primarily this is of concern when either the control or experimental reporter vector, or both, contain very strong promoter/enhancer elements. The occurrence and magnitude of such effects will depend on several factors: i) the combination and activities of the genetic regulatory elements present on the cotransfected vectors; ii) the relative ratio of experimental vector to control vector introduced into the cells; and iii) the cell type transfected. To help ensure independent genetic expression between experimental and control reporter genes, preliminary cotransfection experiments should be performed to optimize both the amount of vector DNA and the ratio of the coreporter vectors added to the transfection mixture. Similar to the firefly luciferase assay, the Renilla luciferase assay is extremely sensitive, providing accurate measurement of ≤10 femtograms of Renilla luciferase, with linearity over seven orders of enzyme concentration. Therefore, it is possible to use relatively small quantities of the pRL-null Vector to provide low-level, constitutive coexpression of Renilla luciferase control activity. The pRL-null Vector, once genetic regulatory domains have been added, can be used for both transient and stable expression of Renilla luciferase. For stable expression, the pRL-null Vector must be cotransfected with an expression vector containing a selectable gene in mammalian cells. Transfection of DNA into mammalian cells may be mediated by cationic lipids (15,16), calcium phosphate (17,18), DEAEDextran (19–21), polybrene-DMSO (22,23), or electroporation (24,25). Transfection systems based on cationic lipid compounds (TransFast Reagent(i), Tfx Reagents(j) and Transfectam Reagent(k)), calcium phosphate and DEAEDextran are available from Promega. For more information and a protocol for the Transfectam Reagent, please request the Transfectam Reagent Technical Bulletin (#TB116) and for the TransFast Reagent, please request the TransFast Transfection Reagent Technical Bulletin (#TB260). Protocols for the use of the Tfx Reagents can be found in the Tfx-10, Tfx-20 and Tfx-50 Reagents for the Transfection of Eukaryotic Cells Technical Bulletin (#TB216). For transfection procedures using calcium phosphate or DEAE-Dextran, please request the ProFection Mammalian Transfection Systems Technical Manual (#TM012).
pRL-null质粒序列载体序列
1 AGATCTCGAG CTCTAAGCTT CACATATGCA TGCACTAGTG GCGCCTGTCG 51 ACGCGTAGAA TTCACCCGGG TACTGCAGAA GTTGGTCGTG AGGCACTGGG 101 CAGGTAAGTA TCAAGGTTAC AAGACAGGTT TAAGGAGACC AATAGAAACT 151 GGGCTTGTCG AGACAGAGAA GACTCTTGCG TTTCTGATAG GCACCTATTG 201 GTCTTACTGA CATCCACTTT GCCTTTCTCT CCACAGGTGT CCACTCCCAG 251 TTCAATTACA GCTCTTAAGG CTAGAGTACT TAATACGACT CACTATAGGC 301 TAGCCACCAT GACTTCGAAA GTTTATGATC CAGAACAAAG GAAACGGATG 351 ATAACTGGTC CGCAGTGGTG GGCCAGATGT AAACAAATGA ATGTTCTTGA 401 TTCATTTATT AATTATTATG ATTCAGAAAA ACATGCAGAA AATGCTGTTA 451 TTTTTTTACA TGGTAACGCG GCCTCTTCTT ATTTATGGCG ACATGTTGTG 501 CCACATATTG AGCCAGTAGC GCGGTGTATT ATACCAGACC TTATTGGTAT 551 GGGCAAATCA GGCAAATCTG GTAATGGTTC TTATAGGTTA CTTGATCATT 601 ACAAATATCT TACTGCATGG TTTGAACTTC TTAATTTACC AAAGAAGATC 651 ATTTTTGTCG GCCATGATTG GGGTGCTTGT TTGGCATTTC ATTATAGCTA 701 TGAGCATCAA GATAAGATCA AAGCAATAGT TCACGCTGAA AGTGTAGTAG 751 ATGTGATTGA ATCATGGGAT GAATGGCCTG ATATTGAAGA AGATATTGCG 801 TTGATCAAAT CTGAAGAAGG AGAAAAAATG GTTTTGGAGA ATAACTTCTT 851 CGTGGAAACC ATGTTGCCAT CAAAAATCAT GAGAAAGTTA GAACCAGAAG 901 AATTTGCAGC ATATCTTGAA CCATTCAAAG AGAAAGGTGA AGTTCGTCGT 951 CCAACATTAT CATGGCCTCG TGAAATCCCG TTAGTAAAAG GTGGTAAACC 1001 TGACGTTGTA CAAATTGTTA GGAATTATAA TGCTTATCTA CGTGCAAGTG 1051 ATGATTTACC AAAAATGTTT ATTGAATCGG ACCCAGGATT CTTTTCCAAT 1101 GCTATTGTTG AAGGTGCCAA GAAGTTTCCT AATACTGAAT TTGTCAAAGT 1151 AAAAGGTCTT CATTTTTCGC AAGAAGATGC ACCTGATGAA ATGGGAAAAT 1201 ATATCAAATC GTTCGTTGAG CGAGTTCTCA AAAATGAACA ATAATTCTAG 1251 AGCGGCCGCT TCGAGCAGAC ATGATAAGAT ACATTGATGA GTTTGGACAA 1301 ACCACAACTA GAATGCAGTG AAAAAAATGC TTTATTTGTG AAATTTGTGA 1351 TGCTATTGCT TTATTTGTAA CCATTATAAG CTGCAATAAA CAAGTTAACA 1401 ACAACAATTG CATTCATTTT ATGTTTCAGG TTCAGGGGGA GGTGTGGGAG 1451 GTTTTTTAAA GCAAGTAAAA CCTCTACAAA TGTGGTAAAA TCGATAAGGA 1501 TCCAGGTGGC ACTTTTCGGG GAAATGTGCG CGGAACCCCT ATTTGTTTAT 1551 TTTTCTAAAT ACATTCAAAT ATGTATCCGC TCATGAGACA ATAACCCTGA 1601 TAAATGCTTC AATAATATTG AAAAAGGAAG AGTATGAGTA TTCAACATTT 1651 CCGTGTCGCC CTTATTCCCT TTTTTGCGGC ATTTTGCCTT CCTGTTTTTG 1701 CTCACCCAGA AACGCTGGTG AAAGTAAAAG ATGCTGAAGA TCAGTTGGGT 1751 GCACGAGTGG GTTACATCGA ACTGGATCTC AACAGCGGTA AGATCCTTGA 1801 GAGTTTTCGC CCCGAAGAAC GTTTTCCAAT GATGAGCACT TTTAAAGTTC 1851 TGCTATGTGG CGCGGTATTA TCCCGTATTG ACGCCGGGCA AGAGCAACTC 1901 GGTCGCCGCA TACACTATTC TCAGAATGAC TTGGTTGAGT ACTCACCAGT 1951 CACAGAAAAG CATCTTACGG ATGGCATGAC AGTAAGAGAA TTATGCAGTG 2001 CTGCCATAAC CATGAGTGAT AACACTGCGG CCAACTTACT TCTGACAACG 2051 ATCGGAGGAC CGAAGGAGCT AACCGCTTTT TTGCACAACA TGGGGGATCA 2101 TGTAACTCGC CTTGATCGTT GGGAACCGGA GCTGAATGAA GCCATACCAA 2151 ACGACGAGCG TGACACCACG ATGCCTGTAG CAATGGCAAC AACGTTGCGC 2201 AAACTATTAA CTGGCGAACT ACTTACTCTA GCTTCCCGGC AACAATTAAT 2251 AGACTGGATG GAGGCGGATA AAGTTGCAGG ACCACTTCTG CGCTCGGCCC 2301 TTCCGGCTGG CTGGTTTATT GCTGATAAAT CTGGAGCCGG TGAGCGTGGG 2351 TCTCGCGGTA TCATTGCAGC ACTGGGGCCA GATGGTAAGC CCTCCCGTAT 2401 CGTAGTTATC TACACGACGG GGAGTCAGGC AACTATGGAT GAACGAAATA 2451 GACAGATCGC TGAGATAGGT GCCTCACTGA TTAAGCATTG GTAACTGTCA 2501 GACCAAGTTT ACTCATATAT ACTTTAGATT GATTTAAAAC TTCATTTTTA 2551 ATTTAAAAGG ATCTAGGTGA AGATCCTTTT TGATAATCTC ATGACCAAAA 2601 TCCCTTAACG TGAGTTTTCG TTCCACTGAG CGTCAGACCC CGTAGAAAAG 2651 ATCAAAGGAT CTTCTTGAGA TCCTTTTTTT CTGCGCGTAA TCTGCTGCTT 2701 GCAAACAAAA AAACCACCGC TACCAGCGGT GGTTTGTTTG CCGGATCAAG 2751 AGCTACCAAC TCTTTTTCCG AAGGTAACTG GCTTCAGCAG AGCGCAGATA 2801 CCAAATACTG TTCTTCTAGT GTAGCCGTAG TTAGGCCACC ACTTCAAGAA 2851 CTCTGTAGCA CCGCCTACAT ACCTCGCTCT GCTAATCCTG TTACCAGTGG 2901 CTGCTGCCAG TGGCGATAAG TCGTGTCTTA CCGGGTTGGA CTCAAGACGA 2951 TAGTTACCGG ATAAGGCGCA GCGGTCGGGC TGAACGGGGG GTTCGTGCAC 3001 ACAGCCCAGC TTGGAGCGAA CGACCTACAC CGAACTGAGA TACCTACAGC 3051 GTGAGCTATG AGAAAGCGCC ACGCTTCCCG AAGGGAGAAA GGCGGACAGG 3101 TATCCGGTAA GCGGCAGGGT CGGAACAGGA GAGCGCACGA GGGAGCTTCC 3151 AGGGGGAAAC GCCTGGTATC TTTATAGTCC TGTCGGGTTT CGCCACCTCT 3201 GACTTGAGCG TCGATTTTTG TGATGCTCGT CAGGGGGGCG GAGCCTATGG 3251 AAAAACGCCA GCAACGCGGC CTTTTTACGG TTCCTGGCCT TTTGCTGGCC 3301 TTTTGCTCAC ATGGCTCGAC

